Observe the directions in which the flowing stripes move.
Slider
Control the animation speed
Button C1
Switch colours from black-white to red-green.
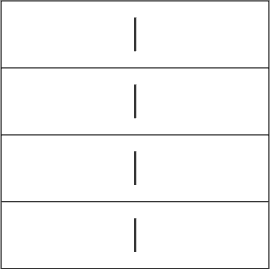
1. When you look at the image and concentrate on grouping the four lines individually, the stripes in these lines start to move vertically only. The direction of movement is upwards in the first and third lines, and downward in the second and fourth.
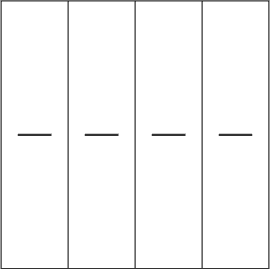
2. When you group the individual columns, they begin to move horizontally only. The stripes in the first and third columns move synchronously to the left, the stripes in the other columns move to the right.
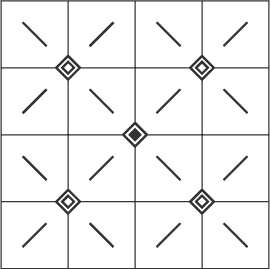
3. When you ignore these previous groupings and look at the image as a whole, you can now see spot-shaped sources near the corners of the image from which black or white diamond shapes constantly emerge. At the centre of the image, there is a hollow, a “black hole” as it were, into which these diamonds then vanish. The direction of movement is now diagonal for all the fields.
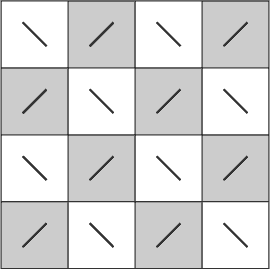
4. You can also group the 16 fields of this kaleidoscope in a chess-board pattern. For example: the second and fourth fields of the first line with the first and third fields of the second line etc. Eight of these fields then create a darker group like a chess-board, which is generally perceived as being in the foreground. The other fields form the lighter background. Which fields are seen as being dark (or somewhat blacker and more distinct) and which as being lighter (or less distinct and veiled in grey) depends on the individual astigmatic deformation of your eyes. If you turn the image or your head around your line of vision, the roles of the dark and light fields usually change. If you cannot see a chess-board pattern, try repeating this fourth experiment with your eyes slightly closed.
When you operate the switch, black and white are replaced by red and green. Make sure you are far enough from the screen and twist it a little until the lightness of the two colours appears to be about the same. (Red and green have different properties of reflection on each computer screen depending on the angle of incidence.)
The diamond shapes that disappear in the centre are normally seen fairly clearly, although they do appear to flow more slowly. The four sources in the corners of the image on the other hand no longer “bubble” as forcefully as in the black-and-white image. At times, these sources seem to dry up altogether. It is now harder to mentally re-group the patterns with the changes in direction of the flow movements that is perceived within the 16 fields. The chess-board pattern – No. 4 - is totally impossible.
In the V1 and upper areas of the visual cortex, there are nerve cells that react to the directions of movement and speed of pixels. Other neural circuits look for corresponding pixels in successive images. The speed of two corresponding pixels and their position in space allows our visual system to determine a local speed with both direction and value. In the MT+ area, we construct entire speed fields. When this is done, not all the local speeds that are registered are taken into consideration. Grouping (in a sequential process between V1 and MT+) creates figure and ground (see Spot 21). This process has a top-down input. You can take control and decide to a large extent how the information from the image is grouped. The distinct points along the outline of a figure are primarily used for constructing the movement made by the figure. Different speeds may be registered at different sections along this outline. These are classified as components in the momentary speed of the figure being constructed (or more accurately, as normal components in the direction of the section of outline in question). A complex figure with relative movement between individual parts is first broken down into object sections (see Spot 21).
The following animation shows an excerpt from the animation at the beginning of the Spot and allows for three different perceptions of movement.
The moving field of stripes is seen as part of a figure which can be observed through the square aperture. The information on how it moves is incomplete since the real outline of the figure is not visible (cf. Spot 20, Attack 5). The movement can therefore be interpreted in a number of ways. For instance: if you start reading the image at the lower edge of the aperture, your visual system registers corresponding points on this edge, and the movement of the partially hidden striped figure is perceived horizontally. However, when you start reading at the vertical edge, you construct a vertical speed for the stripes. Then again, you can start reading at one of the corners and interpret the corresponding points of the two adjacent edges. The two equivalent speed components in this experiment then become combined in a diagonal speed vector for the viewer’s perception. You then have three ways of perceiving what you see. You can trigger top-down input and take control yourself.
The grouping of the parts of the object in the opening animation is governed by top-down input and, as this experiment so impressively shows, occurs before the movement fields are constructed in the MT+ area.
The red-green variant of the opening animation suffers from reduced perception and the image is relatively slow to create for the following reasons: The analysis of periodic structures and complex patterns occurs in the brain’s virtually colour-blind “where channel”, which surrounds the upper visual area in the vicinity of the parietal lobe. The space structured in the foreground, middle ground and background is formed in this channel, together with all available coordinates of the sequentially analysed images in the so-called “what channel”. Processing the red-green image is only possible in this channel to a limited extent due to the lack of contrast in lightness levels. Luckily, the performance of our visual system is redundant. When necessary, the individual images of this animation are processed in the “what channel” which functions slowly and sequentially and which surrounds the visual areas of the two parietal lobes. Peripheral sections of the visual field are neglected because this channel has primarily specialised in interpreting fovea images. One of the places to which the “what and where channels” transmit their findings is the so-called “when channel”, which surrounds the MT+ area and runs above the “where channel”. This is where place and time are linked together and the motion sequences of objects and flowing fields are constructed. The red-green animation is only registered hesitantly because the input of the fast “where channel” is rudimentary in this case, place and time are linked imprecisely, formation of the flowing fields on the periphery is only fragmentary and imperfect. The registered speeds are underrated as a result of the slower construction of the image.
The animation experiment above shows a horizontal rectangular aperture with dominant horizontal edges that supply speeds of corresponding points and thus allow the stripes to be perceived as sliding only horizontally.
It should now be self-evident why the stripes that are arranged identically in all the animations appear to slide in a vertical direction.
In the case of the triangular aperture of the last animation, perception is once again ambivalent
In all three corners of the triangle, the corresponding points are interpreted with their speed components and put together in a variety of ways to create an even speed vector. One of the three measurements is redundant and also contradicts the two other measurements. The assumption of the corresponding points is incorrect, except in the special case with stripes that in reality move precisely in the direction of the edges. The real edge of the striped figure that moves behind the triangular aperture is unfortunately unknown. Lacking this information, our visual system reacts pragmatically and makes do with the measuring points at the edge of the aperture.
The three measurements are evaluated differently. The measurements carry more weight on the edges where the stripes appear to slide slowly and where the distances between corresponding points are therefore short. In order to test this, concentrate deliberately on two of the three edges and examine the various possible speed constructions from two differently weighted speed components. In the case of stripes that slide very quickly the measuring data are generally suppressed completely. The displacement of corresponding points in successive images must not be too great if we wish to allocate the points with subsequent neural interpretation of the speed components.